
By Torsten Liem & Miriam Havel
Summary
This article elucidates the role of the suprachiasmatic nucleus (SCN) in the circadian regulation of the intrinsic temporal program of living organisms and depicts some clinical aspects of its misalignment. Due to its numerous neuronal and neuroendocrine connections, the SCN integrates temporal information of the environment on the one hand and the endogenous biological rhythms of neurohormonal axes – hypothalamic-pituitary-adrenal axis (HPA-axis), hypothalamic-pituitary-gonadal axis (HPG-axis), thyrotropic feedback control etc. – on the other hand. Through these and other complex interaction pathways SCN regulates the neurovegetative system as well as functions of metabolism, immune system and behavior. Clinical aspects relevant for osteopathic treatments are discussed. Keywords Environmental stimulus, circadian rhythm, hypothalamic-pituitary-adrenal axis, hypothalamic-pituitary-gonadal axis, neurohormonal axis, pituitary-thyroid feedback loop, metabolic regulation, autonomic nervous system, neurovegetative system Abstract This article elucidates the role of the suprachiasmatic nucleus (SCN) in the circadian regulation of the intrinsic temporal program of living organisms and depicts some clinical aspects of its misalignment. Due to its numerous neuronal and neuroendocrine connections, the SCN integrates temporal information of the environment on the one hand and the endogenous biological rhythms of neurohormonal axes – hypothalamic-pituitary-adrenal axis (HPA-axis), hypothalamic-pituitary-gonadal axis (HPG-axis), thyrotropic feedback control etc. – on the other hand. Through these and other complex interaction pathways SCN regulates the neurovegetative system as well as functions of metabolism, immune system and behavior. Clinical aspects relevant for osteopathic treatments are discussed. Keywords Environmental stimulus, circadian rhythm, hypothalamic-pituitary-adrenal axis, hypothalamic-pituitary gonadal axis, neurohormonal axis, metabolic regulation, autonomic nervous system, neurovegetative system
The Suprachiasmatic Nucleus (SCN)
Life on Earth has evolved in the context of rhythmic environmental stimuli. The temporal organization in behavior (food intake, sleep, activity) and physiology is common to all organisms and reflects the rhythmicity of the cyclic rotational movement of the sun, moon, and earth. Corresponding to spatial orientation, which requires an internal mapping of the external world, an internal representation of these periods is necessary for meaningful orientation in the temporal space of a day, month, and year. Temporal coordination is controlled by an endogenous temporal program and presupposes a predictable and repetitive rhythm in phylogenesis (Dumbell et al. 2016). In mammals, this endogenous temporal program finds its morphological correlate in the suprachiasmatic nucleus (SCN) of the hypothalamus. This paired group of neurons (in humans, approximately 20,000 densely packed nerve cells) in the anterior hypothalamus regulates the entire internal rhythmicity of the organism as the so-called central circadian pacemaker or “master clock.” In mammals, the internal rhythmicity is hierarchically organized by the SCN, which regulates and synchronizes peripheral oscillators in all other tissues as well as our sense of time through endocrine and neurovegetative signals (Dibner et al. 2010).
Function of the Suprachiasmatic Nucleus
General Function of the Suprachiasmatic Nucleus
Circadian rhythm via light-dark cycles through retinohypothalamic projections (→ pineal gland)
- Waking/Sleeping.
- Circadian-rhythmically controlled activity of the endocrine system, neurovegetative system, behavior (food intake, sleep, activity), metabolism and immune system, body temperature, blood pressure, etc.
The Search for the ClockThe possibility of creating targeted lesions in various brain structures in animal experiments led to the more precise localization of the endogenous pacemaker in mammals. Curtis Richter, who first established the hamster wheel—well known to us in colloquial terms—for the standardized study of spontaneous activity phases of small rodents in the laboratory, determined after numerous meticulous experiments in the 1950s and 60s that rats lost their rhythmic behavior after lesions in the rostral region of the hypothalamus (Richter 1967). Moore and Eichler suspected a close anatomical-functional relationship with the visual system and identified a projection of nerve fibers leading from the optic nerve to the hypothalamus in 1972 using nuclear medicine tracer methods (Moore et al. 1972). At the same time, through experimental lesions of the SCN region in test animals, Stephan and Zucker achieved not only the loss of rhythmic behavior in food intake and the sleep-wake cycle. The ovulation cycle and rhythmic secretion of corticosterone (the most important corticosteroid in rodents) were also disrupted in the otherwise completely healthy animals (Stephan & Zucker 1972). In electrophysiological studies with isolated SCN tissue, a synchronous rhythmic electrical activity of the cells with a phase length of approximately 24 hours could be demonstrated (Inouye et al. 1979). Explantation and transplantation experiments confirmed that the essential mechanism of biological clocks finds expression at the cellular level. The rhythmic spontaneous activity of SCN neurons in vitro and their metabolic activity, as observed in vasopressin secretion and uptake of radiolabeled 2-deoxyglucose, provided further evidence for the central role of the SCN (Schwartz et al. 1980). The more or less accidental discovery of two hamsters in a test animal population that showed a free-running spontaneous sleep-activity rhythm of 22 hours in constant darkness (in contrast to a circadian rhythm of 24.1 hours, as is common for laboratory hamsters) allowed for further clarification of the SCN’s pacemaker function. Biologists Ralph and Menaker succeeded in using fetal SCN transplants from animals with a 22-hour day length to produce this “faster” endogenous sleep-wake rhythm in the “slower” animals. The biological rhythm of the arrhythmic rodents, whose SCN had previously been ablated, was restored and exhibited the circadian properties of the donor (Ralph & Menaker 1988). The central pacemaker function of the SCN, which regulates the endogenous temporal program, was thus clarified. The Path of Light to the SCN The synchronization of the internal clock with temporal cues from the environment is not a passive reaction, but a complex adaptation process (so-called “entrainment”). This temporal-organizational adaptation offers organisms the evolutionary-physiological advantage of anticipating environmental conditions and thus utilizing them optimally. The adaptation process depends on the properties of the endogenous pacemaker and the external stimulus. If temporal information from the outside is missing, as in the hamsters kept in constant darkness by Ralph and Menaker, this leads to the manifestation of the free-running periodicity of the endogenous oscillator (in the case of the “fast” hamsters, these were 22-hour “days”). The external, “entrainment” generating stimulus is referred to as a zeitgeber. Fundamentally, photic (light) and non-photic zeitgebers (e.g., temperature fluctuations, social contact, food intake) can be distinguished. In most living organisms, the zeitgeber is represented by a stable environmental signal of the time of day, the light-dark cycle. For example, the human organism prepares for the start of the day daily from about 4:00 AM. Daylight triggers an entire cascade of physiological changes. The ability of a zeitgeber to adapt a biological system circadially requires that information about the external time of day, month, and year reaches the endogenous oscillator. For most living organisms, (sun)light represents the most important zeitgeber. A close anatomical relationship to the visual system is therefore logical. The histological structure of the eye had already been thoroughly studied for more than a century, and new anatomical findings were not expected when chronobiologists in the mid-1990s provided increasing evidence of another photosensitive system in the eye (besides cones and rods). In an animal model (mouse), they were able to show that visually blind animals with degenerated, non-functional cones and rods continued to present adequate circadian responses to light stimuli; thus, they did not suffer from circadian “blindness.” Upon loss of the entire eye, and thus also the light receptors for the circadian system, no more circadian response to light impulses occurred (Lucas et al. 1999). Light intervention studies in humans also showed that blind subjects with externally intact eyes can be synchronized to the external day with the help of bright light (Czeisler et al. 1995). Although the light reception relevant for the circadian system takes place at the retina, it is not through rods and cones as traditionally assumed. Rather, the path of light leads via specialized, directly light-sensitive retinal cells (so-called intrinsically photosensitive retinal ganglion cells, ipRGCs), which make up about 1% of the total ganglion cell population of the retina and are localized particularly in the inferior-nasal/medial region of the retina (Foster et al. 2002). With their efferent nerve axons, they lead to the SCN via the retinohypothalamic tract (RHT). Morphologically, the RHT is derived from axon collaterals of the axons of the optic nerves, which grow from the optic chiasm into the ventral part of the SCN around the 4th postnatal day (Stephan Zucker 1972). Light detection by the directly light-sensitive ganglion cells of the retina occurs via the photopigment melanopsin, which is phylogenetically much older than the photopigments of the cones and rods (Provencio et al. 2002; Sekaran et al. 2003). The signal transduction cascade leading to the triggering of the cellular action potential is melanopsin-triggered after the photon-absorbing chromophore molecule 11-cis-retinaldehyde is converted into the all-trans state. Initially identified in the retinal ganglion cells of rodents (which are mostly nocturnal), melanopsin could also be isolated in (diurnal) macaques, whose visual system resembles the human one (Dacey et al. 2005). The absorption maximum of melanopsin is at 480 nm, which corresponds to the blue component of the light spectrum. The melanopsin-producing ganglion cells can thus identify the light of dawn (which is particularly rich in blue components) and perceive the transition between day and night due to the changed spectral composition of the light (Foster 2005). In contrast to the visual system, which provides precise information about the spatial and temporal resolution of a light signal (“image forming retinal pathway”), the projection of the circadian system contains information about the intensity and spectrum of the received light (“non-image forming retinal pathway”). These two systems do not work completely separately from each other, as some retinal light information also reaches the SCN from the lateral geniculate nucleus (a brain structure that passes information from the “image forming retinal pathway” to the visual cortex) via the geniculohypothalamic tract, suggesting an interaction (Foster 2005). From the SCN, circadian information is conducted to other brain structures after switching in the paraventricular nucleus and superior cervical ganglion, including the pineal gland, a hormone-producing gland also called the epiphysis. The role of this organ, apart from melatonin production, is not yet clearly understood; a control function of seasonal rhythmic processes, such as reproduction and hibernation, is assumed (Pevet 2000). In reptiles and some bird species, the pineal tissue is directly light-sensitive (hence the term “third eye”) and represents the dominant rhythmic oscillator controlling the daily periodicity (Gaston et al. 1968; Underwood et al. 1990).
Internal Synchronization and Paracrine Mechanisms Within the SCN
Under in vivo conditions, the SCN integrates exogenous and endogenous signals, especially the light-dark cycle, enabling the organism to prepare optimally for environmental conditions. The unique feature of SCN cells is that they maintain their rhythmic activity even without external influences. Autonomous circadian oscillations of neuronal activity have been demonstrated both in explanted SCN tissue and after isolation of individual SCN cells in vitro (Welsh et al. 1995). However, a comparison between SCN tissue cultures and cultures of individual cells shows that the deviations of circadian periods of the cells in brain slices are smaller (Herzog et al. 2004). The individual oscillators, with their quite different endogenous periods, stabilize and synchronize each other. The higher the degree of organization of the cells, the more stable and precise the circadian oscillation. A coherent pacemaker function thus seems to rely on mutual intra- and intercellular processes. The formation of such synchronizing neuronal circuits or neuronal networks within the SCN is achieved through various mechanisms such as cell connections as well as humoral and paracrine factors. From experiments on explanted and isolated SCN cells, it could be concluded that in vivo cell connections of the gap-junction type mediate a significant portion of the synchronization (Long et al. 2005). The neurotransmitters glutamate and γ-aminobutyric acid (GABA) are responsible for local intrinsic signal transmission and are distributed throughout almost the entire SCN (Albus et al. 2005). Light information from the retinohypothalamic tract is also transmitted via glutamate in the ventromedial SCN region, and GABAergic neurons project from the SCN to other regions of the hypothalamus, such as parts of the paraventricular nucleus. In addition to GABA, several neuropeptides, such as vasoactive intestinal peptide (VIP) expressed in the ventrolateral border zone and arginine vasopressin (AVP) produced in the dorsomedial nucleus, are involved in information processing within the SCN and are further involved in the regulation of hormone secretion synchronization of oscillators located peripherally in the tissue (Hastings et al. 2007). Through a combination of pharmacological and genetic manipulations, it was shown in animal models that the circadian rhythm at the cellular level is partly determined by a hierarchically arranged paracrine neuropeptidergic signal transmission. VIP plays a prominent role here, supported by AVP and gastrin-releasing peptide (GRP) (Maywood et al. 2006).
Afferent and Efferent Pathways of the SCN
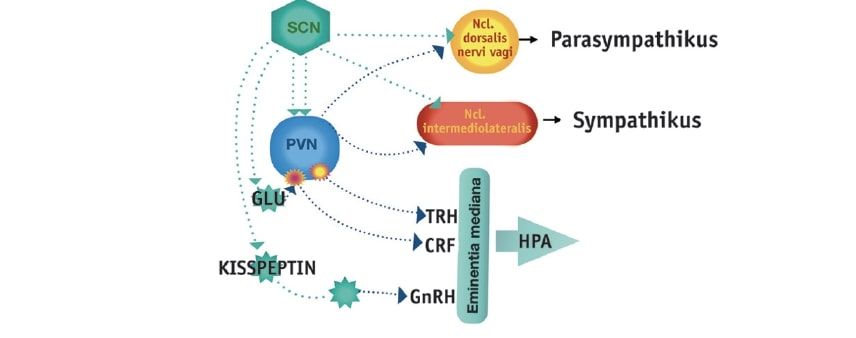
The SCN possesses a multitude of afferent and efferent connections. The aforementioned RHT (retinohypothalamic tract) projects mainly into the ventrolateral region of the SCN, releasing pituitary adenylate cyclase activating peptide (PACAP) and substance P (SP) as neurotransmitters in addition to glutamate, thus transmitting information about ambient brightness for synchronization with the light-dark cycle. Non-photic information reaches the SCN via indirect innervations through the raphe nuclei and the intergeniculate leaflet (IGL), contributing to the synchronization of SCN neurons (Hastings et al. 1996). The core area of the SCN also receives serotonin fibers from the dorsal raphe nucleus and the nucleus raphe magnus of the brainstem. In addition to the mentioned afferent fibers, several other brain regions project to the SCN, e.g., parts of the thalamus, the limbic system, and the hypothalamus (Madeira et al. 2004). Through a variety of direct and indirect efferent projections of the SCN, numerous brain regions are supplied with circadian control signals (Fig. 1). Some connections, especially to large parts of the hypothalamus and the projection to the pineal gland, have been characterized in more detail, which are involved in the control of complex neuroendocrine functions (Abrahamson Moore 2001). Some examples are listed below:Fig. 1: Rhythm-generating neurons of the suprachiasmatic nucleus (SCN) project directly or via interneurons to the paraventricular nucleus (PVN) of the hypothalamus. After switching in the PVN, the projections reach the control centers of the hormonal axes (TRH = thyrotropin-releasing hormone, CRF = corticotropin-releasing factor, GnRH = gonadotropin-releasing hormone) and the autonomous nervous system.
Hypothalamic-Pituitary-Adrenal Axis (HPA Axis)
Corticotropin-releasing factor (CRF) (also called corticotropin-releasing hormone or corticoliberin) producing neurons of the paraventricular nucleus (PVN) of the hypothalamus are innervated by the SCN through direct projections or via interneurons, which causes the circadian release of CRF. In the diurnal state, the arginine vasopressin (AVP) containing axons from the SCN promote the production of CRF and thereby activate the HPA axis (Ulrich-Lai et al. 2006). The circadian control of cortisol secretion via the autonomous SCN-adrenal pathway is controlled by the sympathetic innervation of the adrenal glands (splanchnic nerve). Thus, it could be demonstrated in animal experiments that direct light information is associated with an increased release of cortisol. This occurs independently of the activation of the HPA axis (Fig. 2) (Chung et al. 2011).Fig. 2: Circadian regulation of cortisol secretion and biosynthesis (based on Chung et al. 2011). Fluctuations in circulating cortisol levels are achieved through several regulatory mechanisms: – Modulation of HPA axis activity by the SCN, – Innervation of the splanchnic nerve of the adrenal cortex. Furthermore, it was shown in rats that the circadian release of corticosterone (the most important steroid hormone in rodents) is influenced by modulating the sensitivity of the adrenal glands to ACTH (Ulrich-Lai et al. 2006). The regulation of the sensitivity of the adrenal gland to ACTH stimulation is in turn carried out via the SCN-controlled sympathetic nerve innervation (splanchnic nerve) (Ulrich-Lai et al. 2006). The molecular clockwork of the cortisol-producing cells of the adrenal cortex can also be influenced by the activation of the autonomous SCN-adrenal pathway (Oster et al. 2006; Ishida et al. 2005). Thus, the circadian release of cortisol is subject to direct SCN control through modulation of the HPA axis and through the sympathetic splanchnic innervation of the adrenal gland. However, the local clockwork of the adrenal gland itself also plays an important role, e.g., in maintaining the cortisol rhythm by controlling the capacity and sensitivity of adrenal cortisol secretion and biosynthesis (Chung et al. 2011). Clinical Relevance Dysregulated cortisol secretion can lead to numerous diseases and is associated, among other things, with the development of Alzheimer’s dementia and metabolic syndrome (Carroll et al. 2008; Ferrari et al. 2001; Pasquali et al. 2006; Bao et al. 2008). A decoupling of the fine-tuning between ACTH and cortisol secretion by endogenous or exogenous influences has been linked in some studies to the occurrence of chronic fatigue syndrome, post-traumatic stress disorder, alcoholism, and sepsis (De Kloet et al. 2006). In particular, various potentially disruptive influences such as shift/night work, jet lag, sleep deprivation, or nocturnal food intake can result in serious dysregulations of circadian adaptation in both short-term and chronic cases and lead to the development of various pathologies (Garaulet Madrid 2009; Filipski et al. 2004).
Hypothalamic-Pituitary-Gonadal Axis (HPG Axis)
The SCN influences the gonadotropin-releasing hormone (GnRH) producing neurons in the preoptic nucleus. Furthermore, circadian rhythms interact with peripheral serotonergic systems to regulate lactation (Costa-e-Sousa Hollenberg 2012). The neural oscillator in the SCN is essential for the timing of the preovulatory surge of luteinizing hormone (LH) (Kriegsfeld Silver 2006; De la Iglesia Schwartz 2006). As a correlate for the feedback regulation between the secretion of GnRH and SCN, postsynaptic fibers of GnRH-producing cells could be identified in the SCN in animal experiments (Van der Beek et al. 1997). Lesions of the SCN lead to impaired control of LH secretion, ovulation, and the cycle in female rats (Wiegant Terasawa 1982). Clinical Relevance In the regulation of the HPG axis, it also becomes clear that processes such as ovulation do not represent purely linear top-down processes, but are timed by multi-oscillatory systems and depend on the normal functioning of the synchronization of their coupled components (Sellix Menaker 2010). It is also clinically relevant that disruptions of the circadian timing of the HPG axis, e.g., in night/shift work or intercontinental flights, are involved in the development of infertility, menstrual disorders, menstrual pain, altered duration of the follicular phase, altered FSH levels and follicular phases, low birth weights, and an increased incidence of miscarriages (Fig. 3) (Mahoney 2010; Shechter Boivin 2010). Fig. 3: Hypothesis on a disturbed synchronization of the HPG axis based on Sellix et al. (2010). Circadian oscillators are located in every component of the HPG axis. Disturbed timing of physiological processes depends on desynchronized relationship structures between the SCN, GnRH neurons, pituitary cells, and ovarian cells. Disturbed synchronization in the HPG axis can lead to desynchronization in the HPG axis through a reduction in the amplitude or strength of the circadian clocks in target organs or a change in the amplitude or temporal structures of central or peripheral origin. These can in turn result in disorders of the reproductive organs and reinforce or even trigger reduced fertility (Abbreviations: GnRH gonadotropin-releasing hormone; LH luteinizing hormone)
Thyrotropic Feedback Loop (Hypothalamic-Pituitary-Thyroid Axis, HPT Axis)
The anterior part of the hypothalamus influences circadian variations of circulating thyrotropin (TSH) and thus the circadian fluctuations of peripheral thyroid hormones. This occurs via the projection of SCN neurons into the parvocellular neurons of the paraventricular nucleus (Kalsbeek et al. 2000; Costa-e-Sousa Hollenberg 2012). However, thyroid hormones do not influence circadian clock genes, so the circadian rhythm reacts independently of negative feedback mechanisms, such as the inhibitory effect of free T4 on the secretion of the control hormone TSH (Caria et al. 2009). Possibly, the circadian regulation of the HPT axis is under the control of neuronal pathways, independent of negative feedback mechanisms of thyroid hormones. However, the possibility cannot be excluded that non-genomic processes of thyroid hormones control the excitability of neurons that form the hypothalamic circuit and determine the rhythmicity of TRH release (Costa-e-Sousa Hollenberg 2012). Clinical Relevance Clinically, the destruction of the SCN influences the rhythmic fluctuation of circulating corticosteroids, which in turn exert an inhibitory influence on TRH release (Alkemade et al. 2005). Accordingly, the HPA axis has an inhibitory effect on the HPT axis.
Neurovegetative System (ANS)
Parasympathetic activity is regulated circadially via SCN projections to the dorsal nucleus of the vagus nerve. Sympathetic preganglionic neurons are influenced via connections to interneurons in the thoracic segment. In addition, the neurovegetative system is influenced via the SCN through the innervation of the preautonomic parvocellular neurons of the paraventricular nucleus. Axons located dorsally and caudally in the paraventricular nucleus lead either directly—or via catecholaminergic neurons of the reticular formation—to the intermediolateral nucleus of the thoracic spinal cord section and can thus influence the sympathetic preganglionic neurons, e.g., in the superior cervical ganglion. Light stimulation immediately increases the adrenaline level in the plasma (Ishida et al. 2005). As already explained, the circadian influence of the SCN on the HPA axis is mediated via the neurovegetative innervation of the adrenal glands (splanchnic nerve). Thus, light stimulation leads to an SCN-dependent rapid induction of Per1 expression in the adrenal gland (Ishida et al. 2005).
Pineal Gland (Epiphysis, Corpus Pineale, Pineal Organ)
The circadian control of the pineal gland via efferent pathways of the SCN is comparatively well studied. The endogenous oscillation generated in the SCN is modulated by RHT-mediated information about ambient brightness and projected to the paraventricular nucleus (PVN) of the hypothalamus. During the day, GABAergic inhibition of the PVN neurons occurs, whereas at night, glutamate-mediated excitation of the same occurs (Perreau-Lenz et al. 2003). From the PVN, the signal is transmitted via the intermediolateral nucleus (IML) of the spinal cord and the superior cervical ganglion (SCG) to the pineal gland. Synthesis and release of the pineal hormone melatonin occur at a high rate at night and a low rate during the day. In in vitro studies on brain slices, melatonin receptors were identified in the SCN (Vanecek et al. 1987), which enables direct feedback of the released hormone on the circadian system and consecutively the shifting of sleep-wake phases (Lewy et al. 1980). The nocturnal melatonin signal provides the time information for all cells and organs and is, according to Blask (2009), the most stable and reliable peripheral biomarker of time measurement of the central biological clock (Fig. 4). Fig. 4: Circadian control of the pineal gland: The neuronal pathway includes the paraventricular nucleus (PVN), intermediolateral nucleus (IML), the superior cervical ganglion (SCG), and the pineal gland. Sleep homeostasis depends on sleep pressure and the circadian rhythm. The ventrolateral preoptic nucleus (VLPO) of the anterior hypothalamus is a key factor for falling and staying asleep, while the lateral-posterior part of the hypothalamus is partly responsible for waking behavior (Saito et al. 2018). A narrowly defined area of the lateral-posterior part of the hypothalamus contains specialized neurons synthesizing the neuropeptide orexin, which, in addition to its essential importance for sleep-wake regulation, is also attributed an important role in energy homeostasis and appetite formation. The orexinergic pathways of the lateral-posterior part of the hypothalamus project to large parts of the brain, including the SCN, with most fibers ending in the area near the SCN (Sakurai 2007). In the SCN itself, isolated orexinergic fibers as well as a small amount of orexin secretion and orexin receptors could be identified (Mondal et al. 1999). The latter were detected particularly on cells expressing AVP (arginine vasopressin) and VIP (vasointestinal polypeptide) (Bäckberg et al. 2002). The AVP- and VIP-expressing pathways in turn project to the orexinergic neurons in the lateral-posterior part of the hypothalamus (Abrahamson Moore 2001), which suggests a functional link between the lateral-posterior part of the hypothalamus and the circadian network of the SCN in addition to a close morphological relationship. This finds expression in the circadian fluctuating orexin level in the cerebrospinal fluid (Grady et al. 2006). There is also practical relevance to the suspicion that wearing sunglasses in the morning, especially with orange-colored lenses that block blue light, hinders the resynchronization of the biological clock by sunlight (Sasseville et al. 2006).
Clocks Everywhere: The Organism’s Circadian Clock Network
As explained in the discussion on the circadian regulation of neurohormonal axes, while the SCN can be seen as the central pacemaker, local oscillatory systems at the organ or tissue level also play an important role in circadian regulation. These locally effective circadian mechanisms have been extensively studied experimentally. But even without external light information and social influences, circadian rhythms persist (Czeisler Klerman 1999). In animal experiments, it was shown that circadian clocks occur in practically every cell and every organ tissue and are synchronized by the SCN under normal conditions (Buijs Kalsbeek 2001; Balsalobre 2002). In the event of a time shift and resulting desynchronization between the internal and external time structure, the central pacemaker adjusts comparatively quickly (Yamazaki et al. 2000). However, the peripheral clocks of the internal organs each adapt to the new temporal framework at different speeds and to different extents, which leads to the decoupling of individual rhythms in the circadian network of the organism with resulting symptoms (Hastings et al. 2003). Another aspect of the complexity of the endogenous temporal program lies in the different response behavior of central and peripheral clocks regarding external zeitgeber stimuli. Thus, the SCN reacts to light, whereas the internal clock of the liver reacts primarily to food intake: like other peripheral oscillators, it is only indirectly subject to the influence of light under SCN control (Damiola et al. 2000; Stokkan et al. 2001). Peripheral internal clocks have existed for over 600 million years. Even fruit flies have them. Every cell and every organ has its own clocks; for example, 3–20% of genes exhibit rhythmic expression. Twelve different clock genes and about 20 modulating genes have been identified in humans so far. In the search for the genetic basis of biological clocks, a clock gene of the fruit fly Drosophila was first found. If the molecular mechanism of the intrinsic oscillator was disrupted by a targeted intervention in the fly’s genetic material, it presented arrhythmic activity phases, or these were completely absent (Konopka Benzer 1971). In the 1990s, the first clock gene in mammals (mouse) was identified. Numerous other genes and gene products followed, whose circadian expression reacts to zeitgeber stimuli (Honma et al. 2004; Shearman et al. 2000). According to the current genetic model of the mammalian internal clock, interlocking transcriptional-translational feedback loops (TTL) form the basis for circadian oscillation at the cellular level (Brown et al. 2012). Briefly summarized, during the day, the transcription factor CLOCK (circadian locomotor output cycles kaput) or NPAS2 (neuronal PAS domain-containing protein 2) is bound with BMAL1 (brain and muscle aryl hydrocarbon receptor nuclear translocator-like 1) to E-box (enhancer box) promoters to stimulate the expression of Period (Per1–3) and Cryptochrome (Cry1/2) and others. Thus, over the course of the day, there is an increase in PER/CRY protein complexes in the cytoplasm, which later move into the cell nucleus, where they inhibit the activity of CLOCK-BMAL1 complexes (or NPAS2–BMAL1 complexes). This leads to a shutdown of Per/Cry transcription during the night. Towards the next morning, with the increasing degradation of the PER/CRY complexes located in the cell nucleus, a new cycle begins with an increase in PER/CRY protein complexes in the cytoplasm (Dumbell Matveeva 2016). In addition to visual cells, explicit circadian genes (PER2, 3 or CLOCK) are associated, among other things, with the regulation of cell aging and uncontrolled cell growth (Fu Lee 2003). For example, a defect in the period gene leads to the loss of peripheral rhythm. A molecular biological deletion of a single gene controlling the daily rhythm (PER2) in animal experiments causes the affected animals to age and die dramatically faster than the comparison group of genetically identical rats with intact rhythm genes (Fu Lee 2003; Lee 2006). Peripheral circadian rhythms exist, for example, in the gene expression of the liver, adipose tissue, muscles, mammary gland, and heart tissue (Green et al. 2008). These internal clocks coordinate with each other and show a cycle of 24.20 hours in humans, but are relatively inaccurate compared to the SCN. Through the continuous comparison of cell-internal time measurements and the resulting possibility of self-correction of gene and protein expression with external zeitgebers, especially light information via the SCN, but also through social factors, a precise rhythmic coordination arises in the organism (Pittendrigh 1993). The partial flexibility of the self-excited biological oscillator allows, on the one hand, a harmonious temporal coordination of the physiology and behavior of an organism with its externally or internally conditioned changes. On the other hand, it must remain stably synchronized with the external day in order to be able to anticipate periodic processes in the environment. Every observed circadian rhythm is the product of endogenous and exogenous influences (Fig. 5).Fig. 5: Synchronization of circadian time control in the body. (Modified based on Hastings et al. 2007). Circadian coordination within the organism, in which the primary pacemaker of the SCN, guided by solar radiation from retinal afferents, is maintained and synchronized by tissue-based clocks in the main organ systems and by interrelationships of endocrine, neurovegetative, behavioral (nutritional), and immunological signals.
Clinical Relevance of Findings on Circadian Rhythm
Chronically ill individuals frequently appear to exhibit shifts in their circadian rhythms (Kanikowska et al. 2015; Moore-Ede et al. 1983; Luce 1970). For instance, alterations in sleep cycles impair the immune system (Bollinger et al. 2010). The clinical manifestation of circadian rhythm shifts could therefore be taken into account in osteopathic treatment, particularly in chronic diseases. However, circadian considerations also play a significant role in the therapeutic support of healthy individuals, such as athletes, regarding sleep-wake behavior, jet lag during international competitions, timing of food intake, training routines, and types of exercise (Drust et al. 2005; Hammouda et al. 2013). Differences in time-dependent peak performance can be attributed to internal physiological mechanisms and circadian influences (Facer-Childs Brandstaetter 2015). For example, jet lag caused by traveling across time zones differing by more than 5 hours increases susceptibility to illness by a factor of 2–3 (Schwellnus et al. 2012). Many individuals, especially the late chronotype (the so-called owl phenotype), experience daily recurring circadian disruptions that can affect performance, health, and well-being (Horne Moseley 2011; Roenneberg et al. 2013; Samuels 2009). Conclusion Living organisms exhibit an endogenous periodicity in behavior (sleep and wakefulness, rest and activity, alertness) and physiology (body temperature, hormone release, metabolic processes, etc.). They do not merely adapt to environmental changes reflexively but anticipate them to prepare optimally. The ability to anticipate environmental conditions and processes represents an essential mechanism for the successful survival of a living being and led to the development of endogenous temporal programs over the course of evolution. In mammals, the morphological correlate of the endogenous temporal program is the suprachiasmatic nucleus (SCN) in the anterior hypothalamus. The intrinsic molecular clockwork generates a self-sustaining oscillation with a period of approximately one day (circadian) and is synchronised by stable external environmental stimuli (so-called zeitgebers). The most important zeitgeber for living organisms is (sun)light, i.e. the day–night cycle. The SCN receives information about the intensity and spectral composition of incoming light via specialised retinal ganglion cells and the retinohypothalamic tract (RHT). The neurons of the SCN are synchronised with one another via paracrine mechanisms and transmit circadian information to central and peripheral effectors. In this way, the circadian network in the SCN integrates and regulates both external environmental stimuli and the internal temporal signals of peripheral organ and tissue oscillators. Chronically ill individuals in particular often show shifts in their biological rhythms. Taking these chronobiological shifts into account is an important aspect in the treatment of chronic diseases. In the osteopathic context, this is less about palpable rhythmic phenomena, as has been speculatively formulated in cranial or visceral aspects of osteopathy. Rather, the focus is on an understanding of polyrhythmic phenomena and interactions. In this sense, symptoms are proactive signs that can be understood as an indication for a change in behaviour. Accordingly, the aim is less, for example, to manipulate brain centres palpatorily, and more to support the patient in learning to perceive, read, and understand chronobiological dysfunctions and desynchronisation between the inner and outer time structure, and to initiate targeted behavioural changes or avoidance behaviour in order to improve homeostasis. How do I recognise, for example, at an early stage—ideally before chronic complaints develop—that my stress axis has been activated, impairing my rhythmic patterns such as sleep, and how can I respond and act appropriately? The aim here is to support the patient’s personal responsibility. Psychoeducation and learning experiences that are meaningful for one’s own life are therefore essential aspects of osteopathic interactions as well. Osteopathic manipulative palpation can support these self-regulatory processes and refine the patient’s own body awareness.
Literature
[1] Abrahamson EE, Moore RY. Suprachiasmatic nucleus in the mouse: retinal innervation, intrinsic organization and efferent projections. Brain Res 2001 916: 172–191[2] Albus H, Vansteensel MJ, Michel S, Block GD, Meijer JH, 2005. A GABAergic mechanism is necessary for coupling dissociable ventral and dorsal regional oscillators within the circadian clock. Curr Biol 2005; 15: 886–893[3] Alkemade A, Unmehopa UA, Wiersinga WM, Swaab DF, Fliers E. Glucocorticoids decrease thyrotropin-releasing hormone messenger ribonucleic acid expression in the paraventricular nucleus of the human hypothalamus. J Clin Endocrinol Metab 2005; 90: 323–327[4] Bäckberg M, Hervieu G, Wilson S, Meister B. Orexin receptor-1 (OX-R1) immunoreactivity in chemically identified neurons of the hypothalamus: focus on orexin targets involved in control of food and water intake. Eur J Neurosc 2002; 15: 315–328[5] Balsalobre A. Clock genes in mammalian peripheral tissues. Cell Tissue Res 200; 309 (1): 193–9[6] Bao AM, Meynen G, Swaab DF, The stress system in depression and neurodegeneration: focus on the human hypothalamus, Brain Res. Rev 2008; 57: 531–553[7] Blask DE. Melatonin, sleep disturbance and cancer risk. Sleep Medicine Reviews. 2009; 13: 257–264[8] Bollinger T, Bollinger A, Oster H et al. Sleep, immunity, and circadian clocks: a mechanistic model. Gerontology 2010; 56 (6): 574–80[9] Brown SA, Kowalska E, Dallmann R. (Re)inventing the circadian feedback loop. Dev Cell 2012; 22 (3): 477–87[10] Buijs RM and A. Kalsbeek. Hypothalamic integration of central and peripheral clocks. Nat Rev Neurosci 2001; 2 (7): 521–6[11] Caria MA, Dratman MB, Kow LM, Mameli O, Pavlides C. Thyroid hormone action: Nongenomic modulation of neuronal excitability in the hippocampus. J Neuroendocrinol 2009; 21: 98–107[12] Carroll T, Raff H, Findling JW. Late-night salivary cortisol measurement in the diagnosis of Cushing’s syndrome. Nat Clin Pract Endocrinol Metab 2008; 4: 344–350[13] Chung S, Son GH, Kim K. Zirkadian rhythm of adrenal glucocorticoid: its regulation and clinical implications. Biochim Biophys Acta 2011; 1812 (5): 581–591[14] Costa-e-Sousa RH, Hollenberg AN. Minireview: The Neural Regulation of the Hypothalamic-Pituitary-Thyroid Axis. Endocrinology 2012; 153 (9): 4128–4135[15] Czeisler CA, Shanahan TL et al. Suppression of melatonin secretion in some blind patients by exposure to bright light. N Engl J Med 1995; 332 (1): 6–11[16] Czeisler CA, Klerman EB. Zirkadian and sleep-dependent regulation of hormone release in humans. Recent Prog Hormone Res 1999; 54: 97–130 (discussion 130–132)[17] Dacey DM et al. Melanopsin-expressing ganglion cells in primate retina signal colour and irradiance and project tot he LGN. Nature 2005; 433, 749–754[18] Damiola, F, N. Le Minh et al. Restricted feeding uncouples circadian oscillators in peripheral tissues from the central pacemaker in the suprachiasmatic nucleus. Genes Dev 2000; 14 (23): 2950–61[19] De Kloet CS, Vermetten E, Geuze E et al. Assessment of HPA-axis function in posttraumatic stress disorder: pharmacological and non-pharmacological challenge tests, a review J Psychiatr. Res 2006; 40: 550–567[20] De la Iglesia HO, Schwartz WJ. Minireview: timely ovulation: circadian regulation of the female hypothalamo-pituitarygonadal axis. Endocrinology 2006; 147: 1148–1153[21] Dibner C, Schibler U, Albrecht U. The mammalian circadian timing system: organization and coordination of central and peripheral clocks. Annu Rev Physiol 2010; 72: 517–49[22] Drust B, Waterhouse J, Atkinson G. Circadian rhythms in sports performance-an update. Chronobiology International 2005; 22 (1): 21–44[23] Dumbell R, Matveeva O, Oster H. Circadian clocks, stress, and immunity. Front Endocrinol 2016; 7: 37[24] Facer-Childs E, Brandstaetter R, The impact of circadian phenotype and time since awakening on diurnal performance in athletes. Curr Biol 2015; 25: 1–5[25] Ferrari E, Casarotti B, Muzzoni B et al. Age-related changes of the adrenal secretory pattern: possible role in pathological brain aging. Brain Res Rev 2001; 37: 294–300[26] Filipski E, Delaunay F, King VM et al. Effects of chronic jet lag on tumor progression in mice. Cancer Res 2004; 64: 7879–7885[27] Foster RG, Hankins MW et al. Non-rod, non-cone photoreception in the vertebrates. Prog Retin Eye Res 2002; 21 (6): 507–27[28] Foster RG. Neurobiology: bright blue times. Nature 2005; 433 (7027): 698–9[29] Fu L, Lee CC. The circadian clock: pacemaker and tumour suppressor. Nat Rev Cancer 2003; 3 (5): 350–361[30] Garaulet M, Madrid JA. Chronobiology, genetics and metabolic syndrome. Curr Opin Lipidol 2009; 20 (2): 127–34[31] Gaston, S, Menaker, M et al. Pineal function: the biological clock in the sparrow? Science 1968; 160 (832): 1125–7[32] Gharbi A, Masmoudi L, Ghorbel S. Time of day effect on soccer-specific field tests in tunisian boy players. Adv Phys Educ 2013; 3, 2: 71–75[33] Grady SP, Nishino S, Czeisler CA, Hepner D, Scammell TE. Diurnal variation in CSF Orexin-A in healthy male subjects. Sleep 2006; 29: 295–297[34] Green CB, Takahashi JS, Bass J. The Meter of Metabolism. Cell 2008; 134 (5): 728–742[35] Hammouda O et al. Time-of-day effects on biochemical responses to soccer-specific endurance in elite Tunisian footballplayers. J Sports Sci 2013; (31) 9: 963–971[36] Hastings MH, Best JD, Ebling FJP, Maywood ES, McNulty S, Schurov I, Selvage D, Sloper P, Smith KL. Entrainment of the circadian clock. Hypothalamic Integrat Circad Rhyth 1996; 111: 147–174[37] Hastings MH, Reddy AB et al. Expression of clock gene products in the suprachiasmatic nucleus in relation to circadian behaviour. Novartis Found Symp 2003; 253: 203–17; discussion 102–9, 218–22, 281–4[38] Hastings M, O’Neill JS, Maywood ES. Zirkadian clocks: regulators of endocrine and metabolic rhythms. J Endocrinol 2007; 195: 187–198[39] Herzog ED, Aton SJ, Numano R, Sakaki Y, Tei H. Temporal precision in the mammalian circadian system: A reliable clock from less reliable neurons. J Biol Rhyth 2004; 19: 35–46[40] Honma S, Nakamura W, Shirakawa T, Honma K Diversity in the circadian periods of single neurons of the rat suprachiasmatic nucleus depends on nuclear structure and intrinsic period. Neurosc Lett 2004; 358: 173–176[41] Horne J, Moseley R. Sudden early-morning awakening impairs immediate tactical planning in a changing ‘emergency’ scenario. J Sleep Res 2011; 20: 275–278[42] Inouye ST, Kawamura H et al. Persistence of circadian rhythmicity in a mammalian hypothalamic ‘island’ containing the suprachiasmatic nucleus. Proc Natl Acad Sci USA 1979; 76 (11): 5962–6[43] Ishida A, Mutoh T, Ueyama T et al. Light activates the adrenal gland: timing of gene expression and glucocorticoid release. Cell Metab 2005; 2: 297–307[44] Kalsbeek A, Fliers E, Franke AN, Wortel J, Buijs RM. Functional connections between the supreachiasmatic nucleus and the thyroid gland as revealed by lesioning and viral tracing techniques in the rat. Endocrinology 2000; 141: 3832–3841[45] Kanikowska D, Sato M, Witowski J. Contribution of daily and seasonal biorhythms to obesity in humans. Int J Biometeorol 2015; 59 (4): 377–384[46] Konopka RJ, Benzer S. Clock mutants of Drosophila melanogaster. Proc Natl Acad Sci USA 1971; 68: 2112–2116[47] Kriegsfeld LJ, Silver R. The regulation of neuroendocrine function: Timing is everything. Horm Behav 2006; 49: 557–574[48] Lee CC. Tumor suppression by the mammalian Period genes. Cancer Causes Control 2006; 17 (4): 525–530[49] Lewy A J, T. A. Wehr et al. Light suppresses melatonin secretion in humans. Science 1980; 210 (4475): 1267–9[50] Long MA, Jutras MJ, Connors BW, Burwell RD. Electrical synapses coordinate activity in the suprachiasmatic nucleus. Nat Neurosci 2005; 8: 61–66[51] Lucas R J, Foster R G et al. Photoentrainment in mammals: a role for cryptochrome? J Biol Rhyth 1999; 14 (1): 4–10[52] Luce GG. Biological rhythms in psychology and medicine. Maryland, NIMH; 1970[53] Madeira MD, Pereira PA, Silva SM, Cadete-Leite A, Paula-Barbosa MM, 2004. Basal forebrain neurons modulate the synthesis and expression of neuropeptides in the rat suprachiasmatic nucleus. Neuroscience 2004; 125: 889–901[54] Mahoney MM. Shift Work, Jet lag, and female reproduction. international J Endocrinol 2010; 813764[55] Maywood ES, Reddy AB, Wong GKY, O’Neill JS, O’Brien JA, McMahon DG, Harmar AJ, Okamura H, Hastings MH. Synchronization and maintenance of timekeeping in suprachiasmatic circadian clock cells by neuropeptidergic signaling. Curr Biol 2006; 16: 599- 605[56] Mondal MS, Nakazato M, Date Y, Murakami N, Hanada R, Sakata T, Matsukura S. Characterization of orexin-A and orexin-B in the microdissected rat brain nuclei and their contents in two obese rat models. Neuroscience Letters 1999; 273: 45–48[57] Moore RY, Eichler V. B et al. Loss of a circadian adrenal corticosterone rhythm following suprachiasmatic lesions in the rat. Brain Res 1972; 42 (1): 201–6[58] Moore-Ede MC, Czeisler CA, Richardson GS. Circadian timekeeping in health and disease. New Engl J Med 1983; 309: 469–536[59] Oster H, Damerow S, Kiessling S et al. The circadian rhythm of glucocorticoids is regulated by a gating mechanism residing in the adrenal cortical clock. Cell Metab 2006; 4: 163–173[60] Panda S, Sato T K et al. Melanopsin (Opn4) requirement for normal light-induced circadian phase shifting. Science 2002; 298 (5601): 2213–2216[61] Pasquali R, Vicennati V, Cacciari M, Pagotto U. The hypothalamic – pituitary – adrenal axis activity in obesity and the metabolic syndrome. Ann NY Acad Sci 2006; 1083: 111–128[62] Perreau-Lenz S, Kalsbeek A, Garidou ML, Wortel J, van der Vliet J, van Heijningen C, Simonneaux V, Pevet P, Buijs RM. Suprachiasmatic control of melatonin synthesis in rats: inhibitory and stimulatory mechanisms. European Journal of Neuroscience 2003; 17: 221–228[63] Pevet P. Melatonin and biological rhythms. Biol Signals Recept 2000; 9 (3–4): 203–212[64] Pittendrigh CS. Temporal organization: reflections of a Darwinian clockwatcher. Annu Rev Physiol 1993; 55: 16–54[65] Provencio, I, Rollag MD et al. Photoreceptive net in the mammalian retina. This mesh of cells may explain how some blind mice can still tell day from night. Nature 2002; 415 (6871): 493[66] Ralph MR, Menaker M. A mutation of the circadian system in golden hamsters. Science 1988; 241 (4870): 1225–7.s[67] Richter CP. Sleep and activity: their relation to the 24-hour clock. Res Publ Assoc Res Nerv Ment Dis 1967; 45: 8–29[68] Roenneberg T, Kantermann T, Juda M et al. Light and the human circadian clock. Handbook Exp Pharmacol 2013; 217: 311–331[69] Saito YC, Maejima T, Nishitani M, Hasegawa E, Yanagawa Y, Mieda M, Sakurai T. Monoamines Inhibit GABAergic Neurons in Ventrolateral Preoptic Area That Make Direct Synaptic Connections to Hypothalamic Arousal Neurons. J Neurosci 2018; 11: 2835–2817[70] Sakurai T. The neural circuit of orexin (hypocretin): maintaining sleep and wakefulness. Nature Reviews Neuroscience 200/, 8: 171–181[71] Samuels C. Sleep, recovery, and performance: the new frontier in high-performance athletics. Phys Med Rehabil Clin North Am 2009; 20: 149–159, ix[72] Sasseville A, Paquet N, Sevigny J, Herbert M. Blue blocker glasses impede the capacity of bright light to suppress melatonin production. J. Pineal Res 2006; 41: 73–78[73] Schwartz WJ, Davidsen LC et al. In vivo metabolic activity of a putative circadian oscillator, the rat suprachiasmatic nucleus. J Comp Neurol 1980; 189 (1): 157–67[74] Schwellnus MP, Derman WE, Jordaan E et al. Elite athletes travelling to international destinations >5 time zone differences from their home country have a 2–3-fold increased risk of illness Br J Sports Med 2012; 46: 816–821[75] Sekaran, S, Foster RG et al. Calcium imaging reveals a network of intrinsically light-sensitive inner-retinal neurons. Curr Biol 2003; 13 (15): 1290–8[76] Sellix MT, Menaker M. Circadian clocks in the ovary. Trends Endocrinol Metab (TEM) 2010; 21 (10): 628–636[77] Shearman LP, Sriram S, Weaver DR, Maywood ES, Chaves I, Zheng BH, Kume K, Lee CC, van der Horst GTJ, Hastings MH, Reppert SM. Interacting molecular loops in the mammalian circadian clock. Science 2000; 288: 1013–1019[78] Shechter A, Boivin DB. Sleep, hormones, and circadian rhythms throughout the menstrual cycle in healthy women and women with premenstrual dysphoric disorder. Int J Endocrinol 2010; 259345[79] Stephan FK, Zucker I,. Circadian rhythms in drinking behavior and locomotor activity of rats are eliminated by hypothalamic lesions. Proc Natl Acad Sci USA 1972; 69 (6): 1583–6[80] Stokkan K A, S. Yamazaki et al. Entrainment of the circadian clock in the liver by feeding. Science 291 (5503): 490–3, Storch K F, O. Lipan et al. Extensive and divergent circadian gene expression in liver and heart. Nature 2001; 417 (6884): 78–83[81] Storch KF, Lipan O et al. Extensive and divergent circadian gene expression in liver and heart. Nature 2002; 417 (6884): 78–83[82] Ulrich-Lai YM, Arnhold MM, Engeland WC. Adrenal splanchnic innervation contributes to the diurnal rhythm of plasma corticosterone in rats by modulating adrenal sensitivity to ACTH, Am J Physiol Regul Integr Comp Physiol 2006; 290: R1128–R1135[83] Underwood, H, R. K. Barrett et al. Melatonin does not link the eyes to the rest of the circadian system in quail: a neural pathway is involved. J Biol Rhythms 1990; 5 (4): 349–61[84] Van der Beek EM, Wiegant VM, van Oudheusden HJC, van der Don HA, van den Hurk R, Buijs RM. Synaptic contacts between gonadotropin-releasing hormone-containing fibers and neurons in the suprachiasmatic nucleus and perichiasmatic area: an anatomical substrate for feedback regulation?. Brain Res 1997; 755 (1): 101–111[85] Vanecek J, Pavlik A, Illnerova H. Hypothalamic melatonin receptor sites revealed by autoradiography. Brain Res 1987; 435: 359–362,[86] Welsh D K, Logothetis DE et al. Individual neurons dissociated from rat suprachiasmatic nucleus express independently phased circadian firing rhythms. Neuron 1995; 14 (4): 697–706[87] Wiegant SJ, Terasawa E. Discrete lesions reveal functional heterogeneity of suprachiasmatic structures in regulation of gonadotropin secretion in the female rat. Neuroendocrinology 1982; 34: 395–404[88] Yamazaki, S, R. Numano et al. Resetting central and peripheral circadian oscillators in transgenic rats. Science 2000; 288 (5466): 682–5







